

Tenho defendido a teoria de que o espermatozoide teria se originado de um agente infectante, através de um processo coevolutivo altamente proveitoso para os seres nele enredados. Sustento que essa inusitada transformação tenha dado origem à reprodução sexual, e a toda a vasta e diversificada rede de criaturas imbricada nesse tortuoso modo de replicação. Creio que a estranheza original da proposta já tenha sido abrandada e que, em certo grau, tendo sido tornada mais palatável – ou muito menos intragável do que já foi –, graças a descrições das possíveis etapas do insólito processo que ocasionou a incorporação do parasita ao ciclo de vida do hospedeiro, originando os seres/colônias que se reproduzem sexualmente. Tais descrições, no entanto, naturalmente, não se ativeram a todos os inúmeros detalhes componentes da bizarra transformação do parasita em espermatozoide, tendo enfocado mais enfaticamente as etapas mais gerais e relevantes do processo.
Nas linhas a seguir, descreverei resumidamente a teoria para, em seguida, me concentrar mais especificamente em um detalhe do surgimento da reprodução sexual, qual seja, a transformação do parasita em espermatozoide.
Recapitulando
A reprodução sexual consiste em um modo aparentemente absurdo de reprodução, dada sua ineficiência extrema decorrente precipuamente do desperdício de metade do investimento reprodutivo que é destinado à produção de machos, desperdício cujos resultados se multiplicam, agigantando-se, geração após geração.
A estrondosa desvantagem seletiva decorrente dessa consequência aritmética foi superada graças à elevação imediata da complexidade dos seres gerados tortuosamente através dessa maneira despropositadamente complicada de reprodução. Ocorreu que a enorme complexidade dos seres gerados sexualmente, muito superior à permitida para organismos assexuais (compelidos a se reproduzirem o mais rápido que consigam) jogou-os, de imediato, em um patamar ecológico inatingível pelos seres até então existentes, onde a inexistência de competição garantiu-lhes a sobrevivência, perpetuando, consequentemente, o exagero barroco do perdulário modo de reprodução. Esse argumento foi exposto mais esmiuçadamente em A História do Mundo – parte VI.
De acordo com a teoria que defendo, a reprodução sexual teria decorrido da coevolução entre um parasita infectante de ovos e seu hospedeiro, durante a qual o pequenino ser teria sofrido drásticas alterações com o fito de se camuflar no interior do hospedeiro.
Primeiramente a linhagem parasita teria se empenhado em reduzir os danos causados por sua permanência no corpo do hospedeiro, buscando mantê-lo vivo até a postura dos ovos, quando ambos parasita e hospedeiro, lançariam seus rebentos.
O empenho na camuflagem e redução de danos ao hospedeiro teria resultado na deposição do genoma do parasita junto ao do hospedeiro, onde permaneceria inerte até receber a mensagem de replicação do hospedeiro, na forma de produção de ovos, quando ele próprio se replicaria gerando a nuvem de parasitas que se dispersariam próximas aos ovos do hospedeiro.
O maior impedimento encontrado por uma linhagem de parasitas para efetuar tal procedimento consiste na recuperação do corpo do parasita após a deposição de seu genoma junto ao do hospedeiro.
Ocorre que as especificações fundamentais para a fabricação do corpo do parasita encontram-se inscritas em seu genoma, sendo ativadas todas as vezes que o parasita gera seus descendentes. Ainda que o maquinário celular de replicação envolva dispositivos extragenômicos, quero dizer, mesmo que a reprodução exija informações adicionais presentes em estruturas externas ao genoma, o parasita pode utilizar em seu lugar as estruturas análogas do hospedeiro, ou depositar eventuais estruturas muito específicas no hospedeiro, simultaneamente à deposição do genoma. Desse modo, a recuperação do corpo do parasita, necessária para a reinfecção de novos hospedeiros, seria conseguida de forma similar à gênese tradicional de sua descendência. Tais considerações sugerem que o parasita possuísse um único cromossomo, o que evitaria complicações decorrentes da seleção e agregação de um conjunto específico de cromossomos para encapsulamento no parasita nascente.
Tem-se assim um parasita capaz de infectar um ovo, misturar seu genoma ao dele e permanecer ali, inerte, até a duplicação da célula, quando o material genético do parasita também se duplica junto ao genoma do hospedeiro, onde permanece à espera de se multiplicar e reencapsular, quando informado da iminência da ovoposição do hospedeiro.
.
O grande salto – o maravilhoso fenômeno mágico resultante na reprodução sexual –, teria decorrido do singelo erro de comandar todo o genoma do conjunto – não apenas o cromossomo parasita –, multiplicando-o conjuntamente para, utilizando o equipamento parasita de segregação, encapsulá-lo, transformando o hospedeiro em um retroparasita!
Resulta desse erro uma invasão do corpo do parasita, na qual o hospedeiro executa exatamente a operação inversa à realizada pelo parasita, inoculando nele seu genoma!

Desse estranho modo de inversão parasítica, forma-se uma criatura única, uma colônia que passa a receber os proveitos da interação recíproca
O resultado é estrondoso, dada a retroalimentação estabelecida com a criação da colônia, na qual as vantagens auferidas pela população de hospedeiros favorecem o parasita, cujas vantagens recebidas favorecem os hospedeiros.
Se supusermos que o investimento na fabricação de um espermatozoide seja de um centésimo do necessário para gerar um ovo – estimativa conservadora que poderia ser estendida a um milésimo –, podemos conceber o surgimento de uma criatura cujo número de descendentes, na primeira geração, é 100 vezes maior que a descendência do indivíduo comum. Esse número se multiplica ao longo das gerações, de modo que na segunda, a descendência do primeiro ser a realizar a reprodução sexual tende a chegar aos 10.000 (dez mil!), chegando a um milhão de descendentes estimados para a terceira geração!
Ressaltemos aqui, o inusitado paradoxo ilustrado pelo parágrafo acima, ao contrastarmos a estrondosa vantagem reprodutiva adquirida pelo primeiro ser a efetuar a reprodução sexual, com as desvantagens usuais gritantes decorrentes desse modo ineficiente de reprodução! O absurdo de tal disparidade se esclarece ao considerarmos o evento como um ato de parasitismo, e não de reprodução. Percebe-se, em tal caso, com naturalidade, números condizentes com os relativos a infecções.
Essa vantagem milhonária obtida pelo primeiro ser a se reproduzir sexualmente seria suficiente para esclarecer o sucesso obtido, de imediato, pelos seres coloniais, retroparasitas, que se multiplicaram sexualmente, e que sobreviviam do mesmo modo que parasitas comuns sobrevivem, mas não foi a única.
Pode-se esperar que os parasitas fossem pouco seletivos na escolha de ovos e tentassem infectar o primeiro que encontrassem, dada a dificuldade de encontrar um ovo no oceano. A falta de seletividade resultaria na diversidade imediata dos seres sexuais, em decorrência da hibridação do infectante com tipos diversos. Pode-se conjecturar que, nos primórdios, cada infecção resultasse em algo que identificaríamos, hoje, como uma nova espécie, gerando uma diversidade sem precedentes.
Os híbridos, portadores do duplo genoma (acrescido ainda do genoma do parasita), tenderiam a uma maior complexidade e a somar as habilidades herdadas de ambos os progenitores – fator estabelecido atualmente como a maior das vantagens decorrentes da reprodução sexual. Note que o aumento de complexidade decorrente da incorporação do genoma de cada criatura infectada propicia um afastamento contínuo do nicho ecológico das criaturas sexuais, cada vez mais distantes ecologicamente de suas ancestrais assexuais, muito mais simples que elas.
.
Uma questão se impõe, aqui, uma dúvida. Todo esse desenvolvimento teria sido disparado por um único passo, o erro original no comando de replicação/encapsulamento do genoma do parasita que ocasionou a incorporação do genoma do hospedeiro ao parasita, constituindo o sistema de retroparasitismo. O erro consistiu na não-segregação do genoma do parasita anteriormente ao encapsulamento de seu genoma, resultando no encapsulamento total do genoma da (agora) colônia.
Pode-se supor, em vista disso, que as primeiras retroinfecções ocorressem do mesmo modo, encapsulando todo o genoma da criatura, o que originaria, sucessivamente, seres diploides (possuidores de um duplo genoma), triploides, tetraploides… , aumentando, a cada geração, o genoma das novas criaturas. Tal ocorrência poderia ser, inicialmente, vantajosa, mas, em muito poucas gerações, resultaria em uma sobrecarga extremamente incômoda e deletéria, para o ser.
Em algum momento, talvez desde o início, a criatura travou a multiplicação do genoma integral da colônia impondo a segregação prévia do genoma infectante.
O caminho mais simples para tal ocorrência – embora não o único –, se daria no caso em que, tanto o parasita quanto o hospedeiro originais possuíssem apenas um cromossomo, cada um. Em tal caso, o erro original de multiplicação e encapsulamento do genoma da colônia poderia ter sido disparado pela eventual fusão dos 2 cromossomos. Essa simples ocorrência que teria sido suficiente para disparar todo o processo. Uma vez fundidos os cromossomos do parasita e do hospedeiro, o processo de segregação e encapsulamento do genoma parasita resultaria no sistema de retroparasitismo descrito acima. Tal ocorrência teria impedido, desde o início, a multiplicação do material genômico resultante nas n-ploidias (ou seja, nesse caso a quantidade de material genético seria mantida constante ao longo das gerações).
E teria sido essa a não muito nobre – e parasítica – origem de todos os machos!
Sobre o equilíbrio entre machos e fêmeas, um circunlóquio conveniente
Uma das mais interessantes e bem estabelecidas conclusões matemáticas aplicadas à biologia é a que impõe a equivalência de investimentos em machos e fêmeas. O investimento em um indivíduo corresponde a todo o custo dos progenitores efetuado até a obtenção de uma criatura adulta, capaz de se reproduzir. Tentarei resumir simplificadamente essa dinâmica que se encontra muito bem apresentada no maravilhoso “O gene egoísta”, livro de Richard Dawkins cuja leitura recomendo entusiasticamente.
Suponha que, por alguma razão qualquer, em um dado momento, exista uma espécie cuja quantidade de um dos gêneros, de machos, por exemplo, seja muito maior que a de fêmeas. Digamos que, em tal espécie, os indivíduos, efetivamente, investem mais em machos que em fêmeas resultando na existência de 10 machos para cada fêmea.
Em tal caso, cada fêmea deixará 10 vezes mais descendentes, em média, que cada macho dessa espécie, dado que cada indivíduo descende de um macho e uma fêmea.
Em decorrência da maior fertilidade das fêmeas que dos machos, linhagens que, por qualquer razão, tendam a gerar mais fêmeas, tenderão a deixar mais descendentes que a média. Isso resulta no aumento das linhagens geradoras fêmeas, e no consequente aumento na população de fêmeas até que a quantidade de machos e fêmeas atinja o equilíbrio.
Por essa razão (esboçada acima de forma muito resumida), a quantidade de machos e fêmeas (ou, mais precisamente, o investimento em machos e fêmeas) tende fortemente ao equilíbrio, favorecendo, constantemente, o gênero que, eventualmente, tenha sido, por qualquer razão, previamente desfavorecido.
A origem das fêmeas
Embora a palavra “fêmea” seja aplicada comumente a indivíduos pertencentes a espécies assexuais, a designação se contrapõe a “macho” e não teria sentido se inexistisse tal oposição.
Sob a descrição acima, a reprodução sexual corresponde a uma forma de parasitismo, sendo essa sua origem. Em tal caso, os machos correspondem aos parasitas, enquanto as fêmeas aos hospedeiros.
.
A reprodução sexual emergiu, de fato, sob a forma de uma relação de parasitismo, regida estrita e exatamente pelas mesmas restrições, percalços e vantagens que as relações de parasitismo. Foi necessária a ocorrência de uma espécie de saturação, na qual a infecção – ou o conjunto delas que, como a gripe atual, se desmembrava em um vasto grupo de infecções aparentadas, mas diferenciadas – tendo se alastrado por um vasto contingente de seres, acabou privada de novos hospedeiros, ainda não infectados.
O período anterior a essa saturação deve ser encarado como uma etapa meramente parasitária na qual uns indivíduos se apossavam do corpo de outros com propósitos autorreplicativos. Foi apenas após a saturação – o alastramento geral da população zumbi infectada –, que a definição de machos e fêmeas começou a se delinear. No primeiro instante, o exercício da feminilidade – na forma da produção de ovos –, consistiu em uma maneira de resistir à infecção, uma vez que, do ponto de vista do hospedeiro, cessar a produção de ovos teria significado a extinção. De qualquer modo, a dinâmica parasita/hospedeiro, também privilegiaria indivíduos que produzissem certa quantidade de ovos, ocorrência que garantiria a sobrevivência de alguns dos parasitas da mesma linhagem. A coincidência dos propósitos não surpreende, mas ilustra o avançado estágio em que a interação simbiôntica da colônia já se encontrava.
Foi a ocorrência de uma saturação, quando praticamente todos os ovos depositados acabavam sendo parasitados (ou fertilizados, conforme o ponto de vista), que efetivamente alçou a relação simbiótica a um modo de reprodução. Anteriormente à saturação, indivíduos pertencentes à população da superespécie enredada em um compartilhamento gênico dependente da infecção parasitária ainda tentavam resistir à infecção, enquanto as vantagens adquiridas através da infecção tornavam-se cada vez mais acentuadas.
.
[O paralelismo entre a situação acima, e a das espécies haplodiploides, como as abelhas, nas quais os ovos fecundados geram fêmeas, enquanto os não-fecundados, postos por fêmeas virgens, geram machos, possuidores de apenas metade dos cromossomos das fêmeas, é mais aparente que real. A estabilidade genômica das abelhas é mantida por serem os machos, e não as fêmeas, os indivíduos haploides].

Lembremos agora o pressuposto simplificador inicial de que o hospedeiro possuísse apenas um cromossomo. A suposição, plausível, facilitou a explicação do mecanismo de segregação do genoma, procedimento necessário para a manutenção da quantidade de material genético que, sem ele, aumentaria a cada geração, em decorrência da acumulação do material genético do novo hospedeiro. Nesse caso, a fusão do cromossomo único do hospedeiro com o do parasita permitia o uso do sistema de segregação utilizado desde sempre pelo parasita para se livrar do material genético de cada novo hospedeiro.
Tal pressuposto me parece bastante adequado, uma vez que simplifica e esclarece a questão. As espécies atuais, no entanto, possuem, normalmente, mais de um cromossomo, garantindo que, de um modo ou outro, em algum momento – que pode ter sido o próprio momento inicial –, um mecanismo de segregação do genoma capaz de dividi-lo ao meio teve que ser implementado.
O genoma parasita resolveu o problema acima, de manutenção da quantidade de material genético, de uma forma simples e direta, mas surpreendente desnecessária e confusa, quando observada sob ponto de vista que não considere a origem do mecanismo, que constitui, hoje, uma espécie de relicto, ou atavismo denunciador de sua origem parasitária.
Divisões celulares usuais são efetuadas após a duplicação do genoma da célula, e da segregação das duas partes idênticas resultantes da duplicação, que desse modo recompõe o genoma de cada célula resultante, de forma idêntica à da célula original. Esse método, denominado mitose, é simples e direto – considerada a complexidade do sistema –, e corresponde ao tipo de processo que seria planejado por um engenheiro.
A formação dos gametas, no entanto, ocorre através de um processo extravagante que consiste na duplicação do genoma da célula e sua posterior divisão em 4 partes, efetuada através de duas divisões consecutivas. O processo, denominado meiose, difere de tudo o que ocorre em outras células; é confuso e entremeado por ocorrências únicas, sem similares em outros processos do organismo.
Aos olhos de um engenheiro, o processo parece introduzir uma confusão desnecessária, um complexo conjunto procedimentos tortuosos que poderiam ser eliminados, simplesmente, efetuando uma divisão celular tendo suprimido a duplicação cromossômica prévia. A constatação surpreendente contrasta com a parcimônia usual dos fenômenos naturais – a predileção da natureza pelos caminhos mais simples e diretos.
A idiossincrasia descrita acima, torna-se natural e compreensível quando observada sob o ponto de vista da transformação coevolutiva dos 2 seres. Sob esse ponto de vista, a meiose resultou da estratégia do parasita de resolver o problema de estabilização do genoma, de manutenção da quantidade de material genético ao longo das gerações, imiscuindo-se na geração das células gaméticas, sobrepondo a elas seu próprio método de segregação genômica resultando nas duas divisões celulares que compõem a meiose, processo redutor do número de cromossomos no qual o parasita readquire sua condição original de haploidia.
Pode-se considerar os seres sexuais como componentes de gerações alternadas, uma delas diploide e comandada fundamentalmente pelo hospedeiro, outra haploide, comandada pelo parasita.
.
Após a saturação, quando virtualmente todos os seres passíveis de infecção já se encontravam infectados, a produção de ovos passou a ser compensadora, passando a imperar a dinâmica de equilíbrio entre o investimento em machos e fêmeas. Originalmente, indivíduos hermafroditas, usufruíam da vantagem de garantir seu sucesso reprodutivo através da autofecundação, enquanto a dinâmica de equilíbrio no investimento entre machos e fêmeas se impunha a cada indivíduo, induzindo-o a dividir equanimemente seu esforço na produção de ovos e espermatozoides.
O surgimento dos gêneros, de seres exclusivamente machos, ou fêmeas, ocorrida posteriormente, permitiu às criaturas a especialização na produção de apenas um dos tipos de gametas. Outra vantagem da exclusividade dos gêneros resulta do impedimento da autofecundação, solução tentadora e vantajosa a curtíssimo prazo, mas impeditiva do usufruto das vantagens decorrentes heterozigose.
A pressão no sentido do desenvolvimento de gêneros especializados, seja na produção de ovos, seja na de espermatozoides – tenho me concentrado no reino animal, tratando apenas de espermatozoides, o mesmo vale para gametas em geral – cresce proporcionalmente à complexidade da criatura.
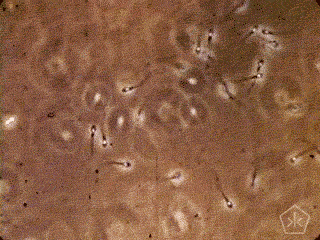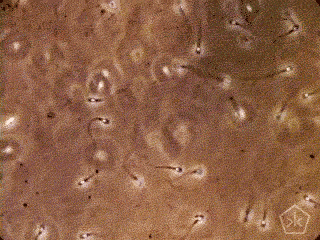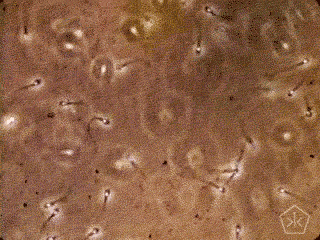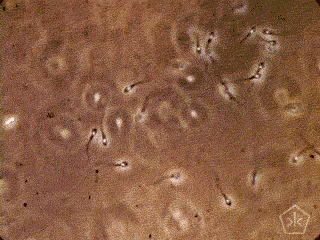
A teoria da origem parasitária do espermatozoide – e da reprodução sexual –, naturalmente, se beneficiará de críticas, necessárias ao acabamento da proposta, algumas delas, apresentadas como objeções a sua aceitação. Gostaria de adiantar uma vantagem da teoria parasitária sobre as outras.
De acordo com a teoria parasitária, a reprodução sexual se originou da tentativa de debelar uma infecção parasitária, fator que acabou resultando, após um longo processo coevolutivo, em um tão intricado entrelaçamento entre parasita e hospedeiro que os transformou em um ser colonial.
O longo processo de incorporação do parasita ao complexo colonial parasita/hospedeiro permite que as objeções à teoria sejam formuladas através de etapas, correspondentes às estratégias de defesa contra a infecção. Teorias que pressuponham o surgimento da reprodução sexual como fator adaptativo puro e simples, no entanto, precisarão responder às mesmas objeções descrevendo um único salto adaptativo capaz de promover todo esse complexo evento.
Veja também: (Os textos sugeridos abaixo tratam exatamente do mesmo tema, cada um deles acrescenta informações que não estão presentes nos outros. Todos eles estão sendo abafados no google – censurados de forma branda –, havendo certa dificuldade em encontrá-los sem o link. A maneira como meus textos sobre evolução têm sido sistematicamente ocultados na internet me faz sentir vivendo no planeta dos macacos).
Gustavo Gollo é multicientista, multiartista, filósofo e profeta.
# Evolução # Biologia # Ciência # Biologia generalizada # espermatozoide # Origem do espermatozoide # reprodução sexual # reprodução sexuada # origem da reprodução sexual # metaevolução # retroparasitismo


Comentários fechados.