

Nicho e interações: Uma introdução à ecologia evolutiva
por Felipe A. P. L. Costa [1]
Competição talvez seja a mais universal das interações e competidores costumam ser agentes seletivos importantes. No caso mais simples, envolvendo duas espécies (A v. B), o sentido das mudanças varia, podendo ampliar ou estreitar a largura do nicho. Vejamos.
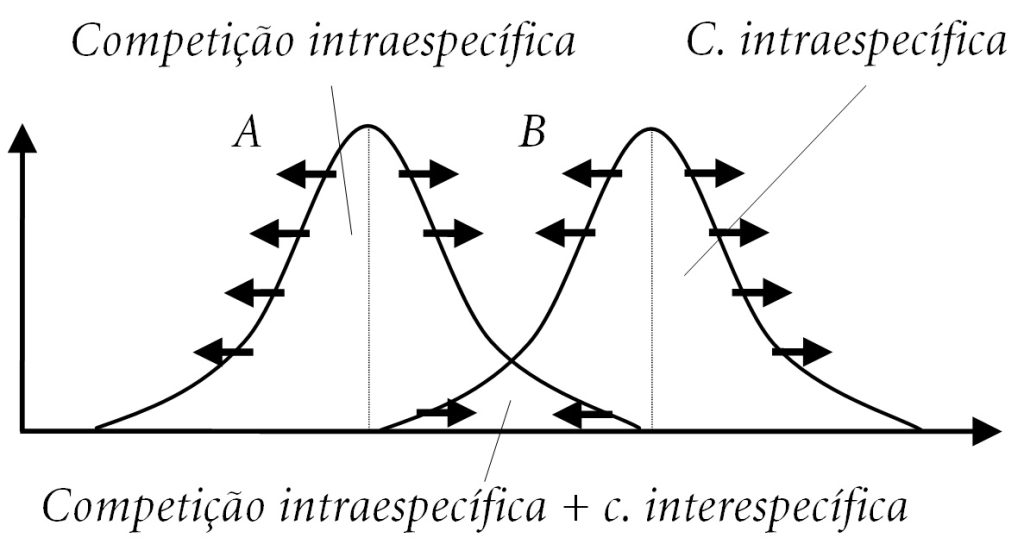
Se a competição intraespecífica predomina (i.e., se A v. A e B v. B são mais intensas do que A v. B), a seleção será centrífuga – tanto em A como em B. Serão favorecidos os fenótipos que exploram recursos extremos (e.g., presas muito menores ou muito maiores que o tamanho médio preferido), em torno dos quais a competição intraespecífica é menos intensa. Isso resultará em divergência intraespecífica, ampliando a largura do nicho. As duas espécies se tornariam mais generalistas, capturando uma maior variedade de presas.
Se a competição interespecífica predomina (i.e., A v. B é mais intensa do que A v. A e B v. B), a seleção tende a ser centrípeta – tanto em A como em B, serão favorecidos os fenótipos que exploram os recursos médios (e.g., presas de tamanho médio, em uma faixa de uso mais ou menos exclusiva). A divergência agora é penalizada. O processo resulta em convergência intraespecífica, estreitando a largura do nicho. As duas espécies deverão se tornar mais especializadas, capturando uma menor variedade de presas.
O nicho, como se vê, não só evolui, mas também está sujeito a ajustes adaptativos [2]. Tornaremos a tratar do assunto.
*
FIGURA. A imagem que acompanha este artigo mostra que os nichos das espécies A e B se sobrepõem parcialmente, indicando competição em uma faixa de uso comum. O impacto evolutivo da situação depende do tipo de competição que predomina: (1) se for a c. intraespecífica, fenótipos que exploram recursos marginais são favorecidos (setas para fora) e os nichos e a faixa de uso comum são ampliados; e (2) se for a c. interespecífica, fenótipos favorecidos são os que exploram recursos intermediários (setas para dentro) e os nichos e a faixa de uso comum são estreitados. (Eixos: e. horizontal, Tamanho da presa; e. vertical, % da dieta.)
*
Interações podem evoluir
Algumas interações interespecíficas são suficientemente íntimas e intensas a ponto de moldar a evolução das linhagens envolvidas. Além disso, a própria natureza da interação pode mudar ao longo do tempo.
Considere o estágio inicial de uma interação entre duas espécies, caracterizado por encontros breves e fortuitos (estágio 0/0). Os encontros podem se tornar sistemáticos, a ponto de emergir alguma interação significativa – e.g., de (0/0) para algo do tipo (0/+) ou (0/–). Dado o primeiro passo, (0/0) → (0/+), digamos, a interação poderá em seguida evoluir para (+/+) ou (–/+). Alternativamente, caso o primeiro passo fosse (0/0) → (0/–), a interação poderia em seguida evoluir para (–/–) ou (+/–).
Vamos dar uma olhada no primeiro caso, (0/0) → (0/+).
A partir de encontros ocasionais (estágio 0/0), duas espécies passam a conviver e a interagir com certa regularidade (e.g., as duas colonizam e prosperam em um mesmo hábitat). Inicialmente, uma delas (dita comensal) apenas se aproveita de restos alimentares deixados pela outra (hospedeiro) – é o estágio (+/0). A relação assim perdura por várias gerações. O aproveitamento dos restos por parte dos comensais é aprimorado e a competição entre eles é intensificada. Nesse contexto, a seleção passaria a favorecer inovações no comportamento de forrageio. Eis então que em um dado momento, além de se aproveitarem dos restos, alguns comensais mutantes passam a pilhar porções do alimento do hospedeiro – estágio (+/–). Emerge então um novo contexto seletivo entre os hospedeiros, no qual toda e qualquer reação de contra-ataque será favorecida – e.g., repelir ou destruir os comensais. Chegamos assim ao estágio (–/–). A história pode terminar aí, com a extinção de uma das linhagens, ou pode prosseguir até um estado de equilíbrio dinâmico – caracterizado por uma sucessão intermitente de ataques e contra-ataques.
Coda
Entender como a evolução mutuamente dependente (i.e., coevolução) tem moldado as interações entre as espécies de uma dada comunidade é um dos maiores desafios da pesquisa ecológica contemporânea. Não é tarefa fácil. Primeiro, em razão da complexidade inerente às comunidades – o quadro geral esboçado nos parágrafos anteriores é uma simplificação. Em sistemas ecológicos reais, as interações significativas costumam envolver não apenas duas, mas várias espécies ao mesmo tempo [3].
Outro complicador é a própria estrutura heterogênea e dinâmica dos sistemas ecológicos. Afinal, as comunidades estão permanentemente sujeitas a mudanças, tanto em escala espacial (“O que tem ali, não tem aqui”) como temporal (“O que ontem era comum, hoje é raro”). A heterogeneidade é traduzida em regimes seletivos flutuantes, conforme tem sido observado em estudos teóricos e de campo – e.g., a identidade e a importância relativa dos agentes seletivos variam de acordo com o hábitat ou a época do ano.
Impostas pela heterogeneidade ambiental, as flutuações no regime seletivo resultam na manutenção dos elevados níveis de variabilidade encontrados em populações naturais. Além disso, ajudam a ampliar e a intensificar a dinâmica da evolução orgânica, fazendo dela a incessante roda-viva a que nos referimos antes [4].
*
Notas
[1] Artigo extraído e adaptado do livro O que é darwinismo (2019), assim como os cinco artigos anteriores – ver aqui, aqui, aqui, aqui e aqui. (A versão impressa contém ilustrações e referências bibliográficas.) Para detalhes e informações adicionais sobre a obra, inclusive sobre o modo de aquisição por via postal, faça contato com o autor pelo endereço [email protected]. Para conhecer outros artigos e livros, ver aqui.
[2] Como o caso das guildas de lagartos em Serra da Mesa GO. Após a extinção de quatro espécies maiores, o lagarto menor que persistiu (Gymnodactylus amarali) passou a ter acesso a um leque mais amplo de presas (cupins). Na ausência daqueles competidores, a largura do nicho (tamanho da presa) foi ampliada, especialmente à direita, visto que houve um aumento na disponibilidade de presas maiores. Os pesquisadores detectaram que, em poucas gerações, o tamanho médio da cabeça dos lagartos aumentou, não apenas em uma, mas em cinco populações de G. amarali – caso único de convergência adaptativa em um sistema que vem evoluindo em tempo real.
[3] A literatura sobre coevolução é copiosa. A adição de espécies torna o sistema bem mais complexo. Considere um predador, P, e duas espécies de presas, A e B. Mesmo sem competir diretamente, A e B podem experimentar competição aparente: a presença de A afeta positivamente P, intensificando a ação e o impacto negativo de P sobre B – e vice-versa. Seria melhor para B (e para A) conviver apenas com P.
[4] Quando os fatores ambientais variam no espaço e/ou no tempo, a seleção pode promover (1) especialização – os fenótipos se tornam adaptados às circunstâncias predominantes; (2) generalização – os fenótipos se tornam adaptados às circunstâncias intermediárias; ou (3) plasticidade fenotípica – a população abriga fenótipos que se ajustam a circunstâncias distintas. Para descobrirmos qual alternativa seria a melhor solução em um caso particular, nós precisamos saber como a aptidão individual é afetada pelas mudanças ambientais. Se a flutuação ambiental extrapola os limites de tolerância, a melhor solução seria maximizar a aptidão em certas circunstâncias (1 e 3); se não extrapola, o melhor é maximizar em circunstâncias intermediárias (2).


Comentários fechados.